 Рис. 1
Рис. 1
Био и Гео: эволюция мыслей Вернадского в мыслях
Айзатуллина
К.М. Хайлов, Институт биологии южных морей НАН Украины,
Севастополь
1. Русский путь в биосферном мышлении
В предисловии к антологии литературы о В.И. Вернадском (В.И. Вернадский: Pro et Contra, 2000), вышедшей, что естественно, в серии "Русский путь", ее главный составитель А.В. Лапо пишет: "Из за границы он (Вернадский - К.Х.) привозит готовую рукопись - это "Биосфера", которая в том же году была издана в Ленинграде. Ею зачитывались Михаил Пришвин, и Николай Заболоцкий, а Геннадий Гор писал: "Есть книги, воздействие которых на читателя ни с чем не сравнимо. Такой необыкновенной книгой оказалась для меня "Биосфера ". В печати тех лет, однако, - продолжает А.В.Лапо - "Биосфера" почти не получила откликов " (с. 16; разрядка моя - К.Х. ).
Эта ремарка, как будто беглая, обозначает, мне кажется, глубинное сходство Вернадского и Айзатулина в характере ума (всеохватность, всюдность, проникновенность) и в жизненном поведении (яркая особливость, нестяжательность, неконформичность). Взятые в совокупности, эти свойства, хоть и с опозданием, на Русском пути отклики получают, а с Западном путем мышления не пересекаются. К нему там глухи. Западный путь прошел мимо биосферы, к бледной Гее.
И Вернадский, и Айзатулин восходили на своем пути в химической плоскости - от масс и концентраций веществ к активностям молекул и к потокам вещества ( косного и живого), а в биогеохимической плоскости -от минералов земной коры, особенно от гидратированных, к геологическим структурам с населяющим их людскими группам, а затем к человечеству. И особо - к России, исторически организованной сложно, противоречиво, конфликтно. Вернадский был и теоретиком, и научно-социальным практиком. Айзатулин, по духу своему - теоретик, стал аналитиком практики российского исторического бытия. Сходство (проследим и дальше) позволяет мне обоих называть- с другими не спутаешь - либо вообще без инициалов, по фамилии, либо только по инициалам: Владимира Ивановича - В.И.. , Тамерлана Афиятовича - Т.А.. Уважение к ним этим умалиться не может.
Часто, слишком часто имя и научный образ Вернадского используются как икона. Но держать ум в раме - нелепость. Ум, учение - как и все природные системы принципиально открыты, растут и развиваются. Закрыть их невозможно (хотя заткнуть их усилием "политической воли" старались и стараются). Концепция биосферы сложилась в первой четверти прошлого века, но ведь не застыла, а росла новыми данным. Во времена Вернадского биокосная (Био-Гео) структура биосферы была почти не ясна, измерений ее параметров было совсем мало. К концу XX века не столько силами самой биогеохимии, сколько тремя научными семействами - науками о Земле, о жизни и о человеческом обществе было накоплено огромное количество численных данных об основных биокосных частях биосферы - водоемах с их биотой, почвах, лесных, луговых, культурных экосистемах разного типа, об особливых (ниже прояснится) поселениях на Земле бактерий, растений, зверей и людей. Подчеркну: накопились сведения не о бактериях, растениях, зверях и людях (это все у биологов и в отделах кадров); накопились сведения именно, о поселениях. Все поселения состоят из двух равноважных, равноглавных частей - Гео и Био. Здесь Гео - это "геподоснова" любой формы живого вещества, термин, не Вернадского, а позднейший, принятый в строительной литературе, но имеющий общебиосферный смысл. Именно общий то смысл Т.А. и заметил.
В терминах науки Вернадский понял биосферу раньше всех и до сих пор лучше всех. Но уже тогда, с порога 20-х годов, он был отвергнут. Биологи 20-80х годов довольны Вернадским быть не могли, что вполне объяснимо: в понятии "живое веществе " неуважительно стирались все специфические признаки организмов разных видов. Вожди (обладатели "политической воли") и философы были еще более задеты. Они считали себя монопольно обладающими знанием человечества, а Вернадский соединил в "живом веществе" биосферы людской мир с миром биологическим. И оказывалось, что о человечестве имеют право судить (ужасно: уж не лучше ли политиков и сопутствующих философов?) люди с совсем другими свойствами - изучающие Землю и Жизнь на ней. Следовать за Вернадским пожелали немногие.
Эволюция мыслей Т.А. почти в точности следовала за идеями В. И. , но не повторяла их иконописно, не рядила в праздные ризы, как на юбилейных собраниях, а серьезно наращивала и развивала. Вернадский - ученик В.В. Докучаева и С.П. Костычева шел от почв и почвоведения. Айзатулин, по сути - ученик Вернадского шел от биокосных систем и знаний другого типа - океанов, морей, озер, от океанологии, лимнологии, лимологии (редкостной науки о границах). Вернадский с суши переходил к гидросфере, сушу, разумеется, не покидая. Айзатулин из океана вышел на сушу, океанов не оставляя. От биоты суши и вод Вернадский перешел к людям, неразрывной части единого, сбалансированного из биогеохимически разных дискретных частей живого и косного вещества биосферы (см. ниже на графиках). Аналогичный шаг, на том же биогеохимическом основании сделал Айзатулин. Вернадский заявил: мысль человека и мыслящее человечество - важнейшая биогеохимическая сила и стал изучать историю мышления (логику и историю науки, сам ею руководя). Историю пристально изучал и Т.А. Вслед за Л.Гумилевым Т.А. обратился к этносам и этнодинамике - крупномсштабным (в географическом пространстве и историческом времени) совокупностям Гео и Социо.
В этносах действует отнюдь не безликая и безбрежная "окружающая среда" биологов. А действует конкретно очерченная, лежащая под ногами именно этой человеческой части живого вещества (не только людей) геологическая основа. Она выступает территориально-обширным (а не частным, например, от отельного мутагенного вещества или от источиника радиации определенного типа) индуктором изменений. Читаем об этом у Т.А. по Л.Гумилеву (Айзатулин, 1999, с. 27 ): " Пассионарный толчек" -микромутация*, вызывающая появление пассионарного признака в популяции и приводящая к возникновению новых этнических систем в затронутых ею регионах. Наблюдается на поверхности Земли в виде полос шириной порядка 200-400 км и длиной примерно 0.5 окружности планеты..."
-------
*) Мне кажется удивительным, что Гумилев выбрал здесь слово "микромутация" Судя по масштабу явления и природе действующих геологических сил, можно, казалось бы, говорить о "макромутациях" или даже о "биогеомутациях". И, ведь, не исключено, что под воздействием сил Земли изменяется не один "признак", а группа признаков изменяется связно.
Прямо как по Вернадскому и Айзатулину, известный молекулярный генетик Лима-де Фариа ( 1991) писал: "Все основные структуры и все основные функции содержат минеральный компонент. который уже был очевиден еще до того, как в общий эволюционный процесс были включены ген и хромосома" (с.41). " Биологическая эволюция существует по той простой причине, что ее нельзя было избежать. Протон, нейтрино, бозон на заре формирования Вселенной обладали качествами, которые делали последующую эволюцию растений и животных неизбежной"(с. 13).
Т.А. сам свой жизненный путь нам раскрыл. Это давний, русский мыслительный путь, пройденный многими думающими, а потому часто из России гонимыми. Системообразующие качества интегральной "русскости" (идущей не от одного национального источника) формируются, по Айзатулину, начиная от химических оснований, но не от химизма макромолекул с вероятностной игрой случайных, частных сил. Качество "русскости" идет от химизма косных географических плит, от биогеохимических провинций, от климатических границ России, зажатой между арктическим безлюдьем севера и пустынями юга. От Земли в географическом смысле начинается многонациональная структура российского населения, его крестьянства (а под ним земля уже как почва), сложносплетенная (мне уже не разобраться.....) историческая этнодинамика.
Биологи полагают, что структуры и функции живого создал естественный отбор. Но на всех уровнях организации функции живого имеют в своей основе потоки химических или физических "частиц" разного размера. При сжатии кристалла кварца в нем возникает поток электронов; и обратным образом: при их протекании через кристалл меняется его форма. Земля с ее мощными силами сжатия и растяжения порождает косные земные формы - географические изложницы. В них ложатся и в них формируются локальные группы живого вещества (не обязательно самих организмов; там, быть может, гуляет генный вихрь и отбор из него; не знаю). С логической неизбежностью Земные изложницы жизни (Гео: косная основа озер, морей, рек, дискретных участков суши) формируют Био - флоры, фауны, биоценозы, этносы, отчасти и государства, а в широком смысле - состав и соотношения масс живого вещества биосферы и его сначала локальные, а затем (этажем выше) глобальные биогеохимические функции - потоки веществ в самих изложницах, к ними и от них. И рассматривают Вернадский и Айзатулин не живых тварей и не некую, без четких границ, "окружающую среду", а в эволюции Земли вычерченные, с измеримыми площадями и объемами, обитаемые биосферные пространства вместе с их населением, т.е. Био-Гео или Гео-Био в зависимости от подхода. Биологам удобнее, чтобы Био и Гео было отдельно. Спору нет, отдельно тоже можно, тоже правильно и хорошо. Но хорошо для других, биологических научных целей. Не хуже и не лучше. Цели Вернадского и Айзатулина - понять обмен веществ, разные проявления биосферной (а не просто организмов) активности, измерить их. Зачем? Да чтобы понять жизнь биосферы, что в задачи биологов не входит по определению (ведь биологическое определение жизни, как формы существования организмов, их видов и сообществ биосферу с ее биокосными частями не предусматривает).
Естественно поэтому, что весь подход к человечеству в биосфере (у Вернадского) и к России в косной части биосферы (у Айзатулина) так различен, непривычен для натурализма классического, дарвиновского типа. Но непривычен - не значит бесплоден. Гораздо более - захватывающе красив, человечен. Потому и читают В.И., открыв "Биосферу", Михаил Пришвин, Николай Заболоцкий и Геннадий Гор.... Потому же и анализ уже свершившейся части этноистории России приводит Т.А. к картинам и интерпретациям новым, неожиданным, хотя нередко и спорным. Ну, и пусть.
2. Продолжая мысли В.И и Т.А.
Биологи изучают организмы, их виды и их сообщества. Численность биологических видов имеет порядок 106. Сообщества биологами не сочтены. Современная экология сконцентрировала свое внимание на экосистемах. Общая численность реально изучаемых экосистем имеют порядок 102-103. Биогеохимия, начиная с Вернадского, рассматривает всю биосферу с человечеством (одна) и крупные ее части, такие, как целые океаны и моря, с их населением (вместе их примерно 150; по спискам в: Морской атлас, 1980). 0бщее число одновременно живущих этносов имеет порядок 101 (сужу по графику в работе: Айзатулин, 1992, рис.1 и 2). Организмы, океаны, моря, разные локальные экосистемы, этносы -важнейшая часть биосферы, но далеко не все разнообразие ее биокосных систем.
Рыбы, моллюски, ракообразные живут в океанах, морях, реках. Но люди содержат их в копаных рыбоводных прудах, в разного рода технических культиваторах, в домашних аквариумах. Птицы селятся в разных местах, но не как попало, а ищут себе подходящего размера, скалу , крону достаточного объема и густоты, болотную кочку с травой такого размера, чтобы гнездо скрывала ( и каждый раз получается особая биокосная система ). Тех же птиц - голубей и кенарей, кур и гусей люди селят в клетки, вольеры. Коров держат в коровниках, овец в овчарнях и так далее. Сами люди живут в биосфере на континентах и островах, но также строят жилые дома, сидят на уроках в школах, в кинотеатрах, в туалетах. Все перечисленное - разные биокосные системы. Большая часть этого списка в работах Вернадского не упоминается. Вероятно как "вненаучные" не приводится они и в учебниках биологи и экологии. Их общая численность вряд ли какой-либо науке известна.
Самую приблизительную оценку численности "вненаучных" биокосных зоосистем можно дать, приняв, что средняя численность обитателей гнезд, нор, различных помещений для скота и других прирученных людьми животных и жилищ самих людей равна 10. В таком случае примерная численность биокосных объектов, не изучаемых ни одной из наук, составила бы 1/10 от числа живущих на Земле особей животных и людей. Это на много порядков превысило бы общую численность всех изучаемых экологией и другими науками групповых объектов с животными.
Заметим у перечисленных выше объектов из числа "вненаучных" то, что заметили у "научных" объектов Вернадский, Гумилев и Айзатулин. Это земная, косная основа ("геопопдоснова"), присущая гнезду пеночки, лисьей норе, скворечнику, конюшне, комнате в общежитии, городскому кварталу. Косная основа здесь не безлика и не безбрежна, как "среда" в биологии. Напротив, это Гео определенного объема с такими-то конкретными свойствами для Био с такими-то конкретными свойствами. Все, как по заказу, адресно. Во всех перечисленных случаях Гео отмерено по потребности обитателей, т.е. соответствует им (их живой массе W) по геометрическому объему (V) , по площади занимаемой земной поверхности (S), по скорости притока ресурсов и оттока собственных, автотоксических продуктов жизнедеятельности и по другим еще многим критериям. Насекомые, птицы, другие звери, люди сами себе значения S и V выбрали. А домашним животным и зверям в зоопарках площадь и обитаемый объем люди назначили по определенной норме. Все численные характеристики меняются от одного биокосного объекта к другому. Может, тогда Гео и Био коррелируют между собой (W с S , We V)? Что коррелировать должны -понятно; вопрос лишь в том, по каким правилам. По общим для всех, или по совсем разным? Правила эти на многих биокосных объектах описаны. Можно их кратко рассмотреть, ссылаясь на имеющиеся публикации.
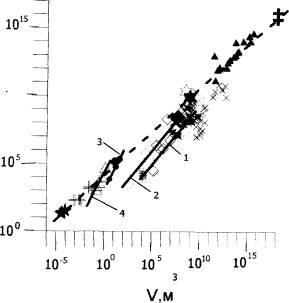
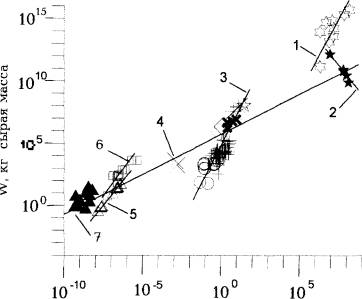
Из двух часто используемых характеристик относительной заполненности обитаемого пространства - W/ S (в биологии - биомасса) и тренда выявляются дискретные группы (1-6) биокосных объектов с высшими и низшими, водными (2-6) и наземными (1) растениями ( для регрессий 1 и 3-6 на рис.2 угловой коэффициент b>1).
Углы наклона регрессий на рис. 1 и 2 являются существенными характеристиками множеств биокосных систем с животными и растениями. Значения углового коэффициента невозможно объяснить биологическими особенности растений и животных, населяющих эти объекты. Напротив, можно с уверенностью сказать, что дискретность групп биокосных систем , определяемая углом наклона соответствующих регрессий, отражает базовые геоморфологические ( V и S) , гидрохимические и почвенные различия, т.е. геоподоснову населяющей их биоты.
Заметим, что в масштабе графика на рис.2 этносы, как один типов биокосных систем, занимали бы положение в крайней правой части шкалы S. Большая часть объектов с растениями и животными простирается на этой шкале до весьма малого размера: S=10' км (экспериментальные модели искусственных рифов: одноклеточные водоросли, поселившиеся в море на керамической основе ). Безотносительно к размеру объекта, свойства его косного ложа (хотя бы только его геометрия; Хайлов и др.,1998) существенны для населяющих его организмов.
Соотношения между Гео и Био в биокосных системах географического ранга (рис.1, 2) являются продуктом природных закономерностей. Люди же строят жилые и рабочие помещения по своим собственным критериям удобства, сами назначают себе и своим хозяйственным животным жилищные нормы (как правило, в терминах W/S). Людские жилищные нормы не имеют в своей основе каких либо теорий. Не вытекают из они и из природных соотношений W(V) или W(S), хотя могли бы. Они являются продуктом тысячелетнего опыта строительства, эмпирического подбора наиболее благоприятных для жизни обитаемых объемов. В связи с этим уместен вопрос: каковы соотношения W и V в жилых комнатах и домах, в рабочих и других помещениях, например, в школах, клубах, кинотеатрах? Тот же вопрос касается и прирученных людьми животных. Отличаются ли соотношения W и V от природных норм в рукотворных биокосных системах - в ульях для пчел, в клетках для кроликов или кур, в овчарнях, в коровниках, конюшнях и в других помещениях?
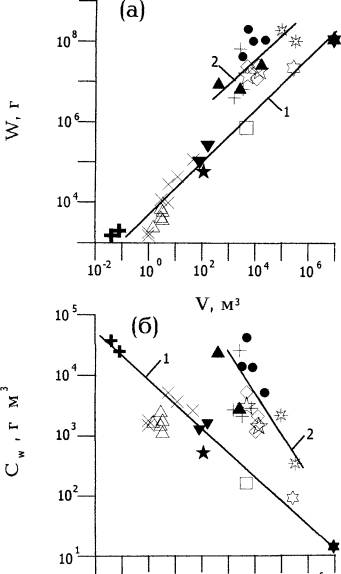
На рис. 3а показано соотношение W и V в жилых помещениях для разных домашних и сельскохозяйственных животных, а также для людей в занимаемых ими помещениях. Как видим, в ряду рукотворных объектов соблюдается правило (1), свойственное природным биокосным системам. Для линии 1 на рис.За: W = 4671*V064 Здесь V - м3 , W - г живой массы.
Совокупность объектов разделилась на две группы. В первую (линия 1)
вошли мелкие одомашненные животные, живущие обычно вблизи людского жилья и используемые
в частном владении (пчелы в ульях, куры и кролики преимущественно в небольших
обиталищах). Сюда же вошли люди в своих квартирах и в городских кварталах.
Первая группа отличается тем, что в нее входят обитаемые объекты разного уровня организации. Высшим уровням здесь являются городские кварталы, состоящие из жилых домов с дворовым пространством между ними. В состав кварталов входят жилые многоэтажные дома. В состав домов входят квартиры. Мелкие животные в небольших клетках нередко являются частью жилых квартир и балконов. Иерархия показанных на графике обиталищ охватывает 9 порядков по занимаемому объему.
Во вторую группу (линия 2 на рис. За) вошли животные среднего и крупного размера, содержащиеся в удаленных от жилья специально построенных помещениях (конюшня, коровники, фермы для крупного рогатого скота), а также люди в общественных помещениях с временным пребыванием (школы, клубы, кинотеатры). Живая масса обитателей во второй группе в среднем на порядок больше, чем в первой при том же объеме помещения. Специфическими чертами второй группы является временное пребывание обитателей в помещениях и (в некоторых случаях) усиленная техническими средствами подача базовых жизненных ресурсов (вентиляция, водопровод, кормовой конвеер в современном коровнике ) с регулярным удалением продуктов жизнедеятельности (канализация) и частой уборкой помещений. Применение разных технических средств, усиливающих обмен веществ в обитаемом пространстве, отчасти объясняют возможность пребывания значительно большей живой массы в помещениях того же объема, что и в первой группе.
Поскольку показатель степени в уравнении (1) для совокупности рукотворных объектов меньше единицы, концентрация живого вещества в обитаемых пространствах снижается с увеличением их объема ( рис.3 б ). В общем виде это соотношение выражает уравнение (2). Для объектов, показанных на рис.3 б (линия 1): b = 0.36 .
Таким образом, на обширном множестве биокосных объектов подтверждается базовое утверждение Вернадского о принадлежности человечества к живому веществу биосферы и утверждение Айзатулина о фундаментальном значении геоподосновы (Гео) для формирования в ее ложе любого живого, в том числе и людского множества (Био-Социо). Дискретные части пространства биосферы с размещаемым в нем строительным материалом жилых и рабочих помещений и живой массой людей или домашних животных - это рукотворные объекты, биогеохимически подобные (по параметрам V и W) природным биокосным системам.
3. Еще один шаг к биосфере по В.И и Т.А.
Объекты, которые Вернадский в одних случаях называл "биокосными телами", в других случаях он называл "естественными телами", а иногда "природными телами". Эти весьма нечеткие определения не могли привлечь серьезного внимания исследователей до тех пор, пока наука не накопила достаточный фонд численных данных, отражающих конкретные характеристики Гео и Био. К концу XX века были выявлены общие для всех биокосных систем численные характеристики, получены их значения и выработаны критерии подобия материальных объектов, каковы бы они не были по генезису. Список "видов" биокосных систем чрезвычайно раздвинулся как в области природных объектов, так и в области их рукотворных аналогов. Вся их известная сегодня совокупность и составляет Био-Гео "начинку" биосферы.
Как разные типы биокосных систем соотносятся между собой в метрическом пространстве биосферы ?
Естественно, что с точки зрения биологии и биогеохимии метрические координаты биосферы и соотношения основных типов объектов представляются существенно разными. Современный взгляд биолога возьмем из работы (Сухонос, 2000, с. 72-73): "...ее (биосферы - К.Х.) масштабный диапазон состоит точно из 15 порядков на шкале десятичных логарифмов (от мельчайших вирусов - 10-5 см до самой биосферы - 109 см), который можно разделить на три одинаковых участка каждый длиной в 5 порядков. Первый участок заполняют одноклеточные системы. Его центр приходится на 10-100 мкм. Второй участок - многоклеточные организмы. Его центр - 1-5 м. Третий участок - популяционные системы. Его центр - 100 -500 км".
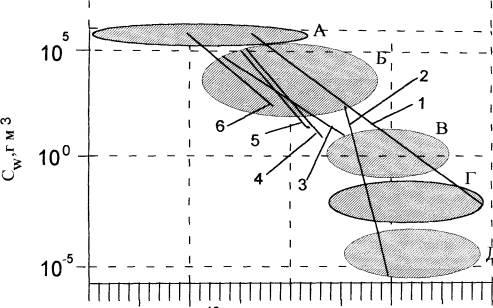
Хотя измерение биосферы и ее частей в единицах длины вполне уместно, интереснее оценить пространство биосферы и ее частей, используя единицы объема. На рис.4 показано взятое из работы (Хайлов, 2001) отображение биосферы с основными группами живого вещества в терминах V и Cw . По обитаемому объему объекты биосферы занимают диапазон от 10-20 до 1020 м3. Поскольку свободные, находящиеся вне живого тела вирусы неактивны, их вряд ли можно относить к числу живых.
Представленные на рисунке 4 биокосные объекты относится к разным ветвям • эволюции - геогенной (водоемы и их геоморфологические части с обитателями), биогенной (растительные пологи, гнезда птиц, норы с их населением) и антропогенной (рукотворные помещения с их поселенцами). Биокосные объекты сформировались в разных средах - морской, пресноводной, воздушной и отчасти почвенной (корневые системы растений, норы почвенных животных). Во всех группах, на разных уровнях организации биосферы, в историческом и индивидуальном времени происходит расширяющийся захват жизненного пространства. Организмы захватывают все большее пространство таким образом, что концентрация живого вещества снижается по определенному закону, отличному от простого физического разбавления. В разных группах биокосных систем тренды Cw по V имеют разные численные характеристики, специфичны. Специфика соотношений Cw и V определяется, по видимому, взаимодействием Гео и Био, а не какой то одной из сторон. Чем больше объем обитаемого пространства, тем мощнее "геоподоснова" жизни в нем, тем (вероятно) большее влияние на нее она оказывает.
По объему индивидуального обитаемого пространства организмы занимают 22 порядка. Концентрация живого вещества в живых телах поддерживается в узком (менее 1 порядка ) диапазоне Cw. Организмы объемом более 2*102 м3 в эволюционно бесперспективны; в биосфере их нет.
Водные биокосные системы надтелесного типа с растениями и животными представлены в биосфере размерным рядом в 30 порядков ( от 10-10 до 1020 м3). В их ряду концентрация живого вещества снижается на 10 порядков ( от 9*105 до 10-6 г/м3 ) .
Вся метрика биосферы охватывает диапазон по меньшей мере в 40 порядков по значениям V и в 12 порядков по значениям Cw. Ранее сообщалось, что интенсивность обмена воды различается в биокосных системах от Мирового океана мельчайших кровеносных сосудов в диапазоне 17 порядков (Хайлов, 1998). Заметим, что логарифмическим центром метрики биосферы является обитаемый объем 1 м3 и концентрация живого вещества 1 г/м3. Примечательно, что величина 1 м3 близка к объему тела человека. Чтобы жить при такой концентрации собственного живого вещества человек с массой 50 кг должен занимать пространство объемом 50000 м3.
В координатах V и Cw условно можно выделить области, отвечающие наиболее важным для человечества жизненным состояниям. Наибольшая концентрация жизни достигнута в телах организмов (на рис.8 - область А), а наименьшая - в глубинах океанов и крупных озер (область Д). В области В располагаются озера среднего размера и высокопродуктивные внутренние моря типа Азовского и Черного. Область Г соответствует большим озерам и морям с меньшей, чем в В концентрацией живого вещества. Наконец, область Б охватывает большую часть рукотворных пользовательских систем (растениеводческих, животноводческих, рыбоводческих). Здесь же размещаются и люди в своих обиталищах. По обитаемому объему логарифмический центр области Б близок к 1 м3, а концентрация живого вещества имеет порядок 103-104 г/м3 (приведенная выше расчетная норма биосферного обитания человека, 5*104 г/м3 , находится вблизи этого диапазона).
Приведенные выше оценки показывают существенное различие метрики биосферы, взятой по биологическим критериям (объекты -организмы и их разные совокупности) и по биогеохимическим критериям ( объекты -биокосные системы, включая организмы, как частный случай). К пространству биосферы биологи обращаются сравнительно редко; подавляющее большинство биоэкологических оценок, начиная с биомассы (W/S ) делается на единицу площади. Концентрацию живого вещества в воде и воздухе экологи рассчитывают, но систематических сравнений по всему разнообразию жизнеобитаемых объектов до удивления мало. В биологической и биогеохимической метрике биосферы существенно (на несколько порядков) различаются ее кардинальные точки (центры).
Однако, указывая на различие двух метрических образов биосферы, подчеркнем их методологическое сходство. Образов биосферы может быть создано много; это зависит от выбора точки зрения и избранных параметров. Разные модели не следует рассматривать в терминах "хуже-лучше"; они взаимно друг друга дополняют. Иное дело, что разные модели отвечают на разные вопросы.
Метрика биосферы, обобщающая основные типы биокосных систем, выполняет познавательную роль аналогичную познавательной роли филетического древа, обобщающего основные таксономические группы организмов. Однако, организменная форма жизни занимает в метрике биосферы довольно скромное место. Оно ограничивает эволюционные интерпретации организменной биологии. Понятно, что в области биокосных систем с ничтожно малой концентрацией живого вещества механизм дарвиновской эволюции, основанный на генетической наследственности и ее изменчивости, творческой роли играть не может. Большая часть объектов биосферы и она сама, как единое целое, могут быть только продуктом коэволюции Земли и жизни на ней. Механизм этой коэволюции, вероятно, не прост и почти не изучен (Лима-де-Фариа, 1991).
Наконец, самое главное: биология, изучающая жизнь особей, видов, и их сообществ, в начале XXI века по прежнему исключает из своего рассмотрения ментальные, трудовые, социальные проявления человека и человечества, а соответственно все их вещественные и духовные продукты, исключает науку, культуру, искусство, мораль, т.е. высшие проявления Человека разумного, самой биологией признаваемого эволюционным венцом жизни. Согласимся, что биология имеет право сама устанавливать свои предметные границы и не включать в научные модели объекты (человечество, косное вещество Земли), за эти границы выходящие. Бесспорно, что в очерченных ею для себя границах заслуги биологии велики. Она учит, как вырастить больше колосьев и больше коров, как бороться с болезнями, как лучше питаться, дольше жить, учит многому еще. Но зато другие группы организмов и их виды живут короче, болеют, вероятно, больше людей, так как их здоровая пища заменяется нездоровой или не в нужном количестве. И не решается главный, коренной вопрос современности - как человечеству следует правильно жить на Земле, в ее биосфере? Заметим, что этот вопрос выходит за пределы биологии на вполне законном основании - в соответствии с биологическим определением Жизни, которое биосферу не предусматривает.
Чтобы ответить на коренной вопрос, другие науки, прежде всего изучающие Землю, постепенно создают более полную научную модель жизни биокосных систем в составе биосферы, включающую и косное вещество, и человечество со всем разнообразием антропных биокосных систем. География на своих картах давно и наглядно соединяет косные, живые дочеловеческие и антропные объекты. Модель биосферы Вернадского делает то же самое, но в гораздо более глубоком и аналитичном виде. На современном этапе своего развития модель биосферы, вобрала в себя научные данные многих дисциплин и обладает разрешающей способностью, адекватной современным задачам человечества, гораздо более сложным, чем в прошлые века/
Одна из актуальнейших задач на пути к выживанию по законам биосферы - переориентация образования из видоцентрической плоскости в биосфероцентрическую. От изучения несомненно важного видового разнообразия жизни давно пора переходить к изучению гораздо более мощного (рис. 1, 7, 8) разнообразия биокосных систем биосферы и ее в целом. На пути к такой переориентации лежат привычные штампы, накопившиеся мифы и другие трудности.
При явно уже обнаруженной зависимости человечества от биосферы,
западная система образования
по-прежнему остается в границах биологического понимания Жизни, как формы
существования организмов, их видов и их сообществ. Но в каком состоянии это
понимание находится и достаточно ли его человечеству? Список столь ценимых
биологических видов Земли далек от завершения и не ясно, когда это произойдет.
Однозначного критерия для оценки размера разных биологических видов (общая ли
численность или общая живая масса особей каждого вида? или общая площадь,
занимаемая видом на поверхности Земли?) не выработано, надежных численных
оценок мало. Поскольку границы групповых объектов чаще всего размыты,
неизвестно ни общее количество сообществ бактерий, растений и животных на
Земле, ни индивидуальные размеры сообществ. А раз нет меры объектам, невозможно
их численное сравнение. Флоры и фауны
выделены по другим основаниям, чем организмы и виды, с ними не сопоставимы. При
всей познавательной ценности филетического древа, как главного отображения
системы Жизни, она не получила
метрики, позволяющей оценивать пропорции Жизни по общенаучным (а не по
специфическим для организмов и их видов) характеристикам. Каков же тогда общий
облик Жизни? Или модели, интегрирующей разные объекты биологии - организмы,
сообщества, флоры, фауны и т.д. не может быть в принципе? Ответа биологи не
дают.
- - - - -
Последний раздел этой памятной статьи от главной ее темы не
отвлекается. Пафос завершающего шага в творческой жизни Тамерлана Афиятовича -
синтез Гео и Социо, как части глобального живого вещества. Эта задача наукой только
лишь начинает осваиваться и Айзатулин останется в числе пионеров.
Предшествующие шаги Т.А. пролегали в Океане, а затем на его освоенных людьми
берегах. Они были не менее значительны.
Это все тот же исконный Русский путь, идущий неведомо откуда, но явно
обозначенный мыслителями-космистами, натуралистами и религиозными философами, а
затем Вернадским. А по Айзатулину (1992,с. 24) - "Судьба России -
судьба ноосферы".
Подрисуночные подписи к статье К.М.Хайлова " Био и Гео : эволюция мыслей Вернадского в мыслях Айзатулина"
Рис.1. Соотношение объема обитаемых пространств (V) и общей живой массы (W) в биокосных системах с животными разных таксономических и экологических групп ( по: Хайлов, 2001).
Группы объектов и соответствующие регрессии: 1 - карпы в прудах (¯) и рыба разных пород водохранилищах (); 2 - зообентос в одесских лиманах (◊); 3 - товарная форель в садках (●); 4 - молодь форели в бассейнах (D).
Отдельные объекты: рапаны из Черного моря (✭), икра лососевых рыб в проточных культиваторах (+) ;зоопланктон и зообентос в озерах (х) ; зообентос в Стенцовских (¨), в Жебрияновских (´) плавнях и в Жебрияновской бухте (✱); зообентос в разных морях (▲); зоопланктон и зообентос в Мировом океане (✢) ; зообентос оз. Донузлав (✲). Штриховая линия - регрессия, обобщающая все морские объекты, з:
Рис.2. Соотношение площади занимаемой поверхности ( S ) и общей живой массы (W) в биокосных фитосистемах с разными таксономическими и экологическими группами растений; по: (Хайлов, 2000 с дополнениями).
1 - разные фитосистемы суши; 2 - разные геоморфологические зоны Мирового океана с фитопланктоном; 3 - водные фитосистемы с разным биологическим составом населения: озерах Польши (✢) и Белоруссии (❍) с сообществами фитопланктона; пологи макроцистиса (✲) и зостеры (✲), разные участки Днепровского водохранилища с зарослями высших водных растений (◇); водохранилища волжского каскада с зарослями высших водных растений (х); 4 - водные растения в небольшом пруду (х); 5 - пологи ламинарии на подводных валунах; 6 - литоральные ванны с донными водорослями; 7 - экспериментальные искусственные рифы с диатомовым обрастанием.
Рис. За. Соотношение объема обитаемых пространств (V) и общей живой массы (W) в биокосных системах с одомашненными животными и людьми.
Линия 1: пчелы в ульях (+), кролики в клетках (△), куры в курятниках (X), двухкомнатная квартира с одним жильцом (☆), однокомнатная квартира с двумя жильцами и трехкомнатная с пятью жильцами (▼) , птичник на 500 кур (□), городской квартал из 2-х 9-этажных домов вор дворе площадью 100*100 м (✱), городской квартал из 20 9-этажных домов с общим населением 2500 человек. (✱),
Линия 2: свинарник (+), фермы для крупного рогатого скота (*), коровники (●), кинотеатр (▲), сельские клубы (☆), школы (◊).
Рис.Зб. Соотношение объема обитаемых пространств (V) и объемной концентрации живой массы (Cw) в биокосных системах с одомашненными животными и людьми. Обозначения как на рис. За.
Рис.4.0сновные группы живого вещества в метрике биосферы (по: Хайлов, 2001):
Линии: 1 - разные морские зоосистемы; 2 - зоопланктон и зообентос в озерах;
3 - пологи высших наземных растений; 4 - пологи морских макрофитов;
5 - кроны фукусов и аскофиллумов; 6 - пологи морских многоклеточных и одноклеточных водорослей на поверхности искусственных рифов.
Овалы: А - область, занимаемая в метрике Cw(V) телами живых организмов -от одноклеточных водорослей до крупнейших животных; Б - область, занимаемая системами интенсивного сельскохозяйственного растениеводства, животноводства, морской фитоаквакультурой, жилыми и прочими человекообитаемыми системами; В - область рыболовства в озерах и малых морях; Г- область крупных озер, морей и океанических вод с низкой концентрацией живого вещества; Д - область глубинных океанических вод и больших олиготрофных озер.
Литература
Айзатулин Т.А. Судьба России - судьба ноосферы (к естественно-научной
теории динамики России в контексте гео- и этнодинамики) // Alma Mater. Вестник высшей школы.
1992.№7-9. С. 24-34.
Айзатулин Т.А. Теория России. Геоподоснова и моделирование. Серия
"Семинары С.Г. Кара-Мурзы". М.: 1999. 69 с.
Лапо А.В. Миры Вернадского: от кристалла до биосферы // В.И. Вернадский: pro et contra. Антология литературы о В.И. Вернадском за сто лет (1898-1998). Изд. Русского Христианского гуманитарного института. СПб.2000.С.7-28.
Лима-де Фариа. А. Эволюция без отбора. Автоэволюция формы и функции. М.:Мир. 1991. 455с.
Морской атлас. Термины, понятия, справочные таблицы. М.: Мин. обороны.
СССР. Морфлот. 1980. 187 с.
Сухонос С.И. Развитие ноосферы в аспекте основных эволюционных процессов во вселенной // Ресурсы ноосферного движения. Материалы международной конференции. М.: Геос.2000.С. 72-73.
Хайлов К.М. Междисциплинарные вопросы на границе наук о жизни и наук о Земле // Известия АН РАН. Сер. Геогр.,2000. № З. С. 30-37.
Хайлов К.М. Что такое жизнь на Земле? Одесса.: 2001.149 с.
Хайлов К.М., Юрченко Ю.Ю., Смолев Д.М., Празукин А.В. Геометрические
условия заполнения гидробионтами пространств и поверхностей их искусственных
жилищ // Успехи совр. биол.1998. T.I 18.
Вып.5. С. 585-596.
Рис. 1
S,
км2
Рис. 2
10-2 10° 102
104 106
V, м3
Рис.
3
10-20 10-15 10-10 Ю-5 100 105 1010 1015 1020
V,
м3
Рис. 4.